In-situ observation of the "dislocations" and "inclusions" inside protein crystals during growth by advanced optical microscopy;
光学顕微法を用いた,成長している最中のタンパク質結晶中の「転位」および「インクルージョン」のその場観察
本ページではレーザー共焦点微分干渉顕微鏡(LCM-DIM),偏光顕微鏡(BM: birefringence microscopy),および位相差顕微鏡(PCM: phase contrast microscope)を用いると,成長している最中のタンパク質 結晶中の「転位」や「インクルージョン」を,完全に非接触・非破壊でその場観察できることを紹介させていただきます(G. Sazaki, et al., Crystal Growth and Design, 5, 1729-1735 (2005)).これまで格子欠陥の観察には,溶液を除去する(X線トポグラフ法の場合),表面をチップでスキャンする(原子間力顕微鏡の場合),溶解させる(エッチング法の場合)など,結晶を通常の成長中とは異なる条件下におく必要がありましたが,本手法は,結晶が成長しているそのままの条件で,非接触・非破壊に観察出来るところに最大の特徴があります.
1)高品質なタンパク質結晶とは何か,2)格子欠陥のその場観察法に望まれる特質と本研究で用いる光学顕微法について,
3)転位(LCM-DIM)およびインクルージョン(BM)のその場観察,4)3)での像が転位であることの証明(エッチング法による) ,
5)結晶中に観察される2種類の格子欠陥:「転位とマイクロ欠陥」,6)インクルージョンと転位の関係
1)高品質なタンパク質結晶とは何か?
 高品質なタンパク質結晶って,いったん何でしょうか?2つの見方があると思います(右図).構造生物学の方にとっては,やはり高分解能の解析に耐えうるX線(中性子線)回折特性の良い結晶のことをを指すでしょう.しかし,結晶成長学の観点からみると,良い結晶とは,あくまで点欠陥,転位,面欠陥,インクルージョンなどの格子欠陥が少ない結晶のことを指します.現在の最大の問題は,どのような種類の格子欠陥が結晶の回折特性に特に悪影響を及ぼすか,この相関がまだ不明な点にあります.
高品質なタンパク質結晶って,いったん何でしょうか?2つの見方があると思います(右図).構造生物学の方にとっては,やはり高分解能の解析に耐えうるX線(中性子線)回折特性の良い結晶のことをを指すでしょう.しかし,結晶成長学の観点からみると,良い結晶とは,あくまで点欠陥,転位,面欠陥,インクルージョンなどの格子欠陥が少ない結晶のことを指します.現在の最大の問題は,どのような種類の格子欠陥が結晶の回折特性に特に悪影響を及ぼすか,この相関がまだ不明な点にあります.
それでは,どうすればこの問題点にアプローチできるでしょうか? 私は,
1)特定の種類の格子欠陥のみを豊富に含むタンパク質結晶を成長させ,そのX線回折特性を調べる,
以外に方法はないと考えています.そのためには,まずそのようなタンパク質結晶を成長させる必要がありますが,そのためには,
2)各種の格子欠陥が
生成するメカニズムを知る必要があります.そのためには,当然,
3)
タンパク質結晶の「成長中」に,各種格子欠陥を非接触・非破壊でその場観察する必要があります.
そこで,そのような格子欠陥のその場観察手法の開発に取り組みました.
(転位・インクルージョンのその場観察ページのトップへ)
2)格子欠陥のその場観察法に望まれる特質と本研究で用いる光学顕微法について
これまで,結晶中の格子欠陥の種類や分布を観察するために,主に,X線トポグラフ法,原子間力顕微鏡法,エッチング法などが用いられてきました.それぞれの長所・短所を下左図にまとめてみました.それぞれの方法には一長一短があります.そこで,上述の我々の目的を達成するためには,,成長中の結晶内部の格子欠陥を非接触・非破壊でその場観察するための手法を開発する必要があることがおわかりいただけることと思います.
そのような手法としては,やはり光学的な手法が一番有力であると思われます.そこで,これまでに行われてきました中で有力と思われる光学的顕微法を下右図にまとめてみました.レーザー共焦点微分干渉顕微法を用いるとタンパク質結晶の単位成長ステップが観察できますし(G.Sazaki, et al., J. Crystal Growth, 262 (2004) 536),偏光顕微鏡を用いて無機結晶中の転位を観察し,その種類を歪場の形状より同定する研究も行われています(K. Maiwa, et al., J. Crystal Growth, 98 (1989) 590).さらに,大きく完全生の良い結晶を成長させることができれば,従来用いられてきた位相差顕微法を用いることでも,タンパク質結晶上の単位成長ステップをその場観察できることがわかっています(K. Tsukamoto, et al., J. Jpn. Assoc. Crystal Growth, 30 (20003) 81).そこで我々は,これらの光学顕微法を用いて,タンパク質結晶中の格子欠陥のその場観察を始めました.


(転位・インクルージョンのその場観察ページのトップへ)
3)転位(LCM-DIM)およびインクルージョン(BM)のその場観察
まず,レーザー共焦点微分干渉顕微鏡(LCM-DIM)と偏光顕微鏡(BM)を用いて,成長している最中のリゾチーム正方晶系結晶{110}表面上に発達した渦巻成長丘をその場観察した例を下図に示します.LCM-DIM法では,結晶表面上の美しい渦巻成長ステップが観察されており,この結晶面が1つの渦巻成長丘で覆われていることがわかります.また,LCM-DIM法,BM法ともに,結晶中のグロース・セクタ・バウンダリーやストリエーションを明瞭に観察できていることがお分かりいただけることと思います.
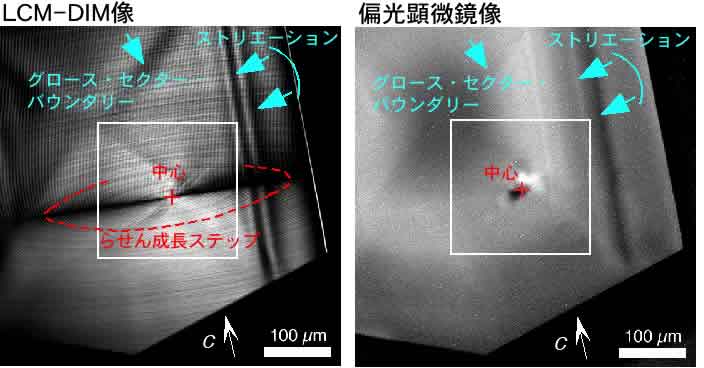
上図において白四角で囲まれた渦巻成長丘の中心部分を拡大した像を下図に示します.LCM-DIM像では,渦巻成長ステップのほかに,赤矢印ln1-ln6で示した「線状」のコントラストが観察できていることがわかります(+印は渦巻成長丘のちょうど中心を示します).これらの線状のコントラストは,後ほど証明しますように,光軸と垂直平面内に発達した「転位」を表しています.
また,同じ部分を観察したBM像では,渦巻成長丘中心部分に「白および黒」のコントラストが観察されます.試料を消光位から反対方向に回転させると,白→黒,黒→白というようにコントラストの白黒が反転します.このことより,これらのコントラストは互いに逆符号の複屈折を有する歪場であることが分かりました(それぞれ引っ張り歪みと圧縮歪みに相当).後ほど証明しますように,これらは渦巻成長丘直下にある固相インクルージョン(微結晶やゴミなど)を表しています.

(転位・インクルージョンのその場観察ページのトップへ)
4)3)での像が転位であることの証明(エッチング法による)
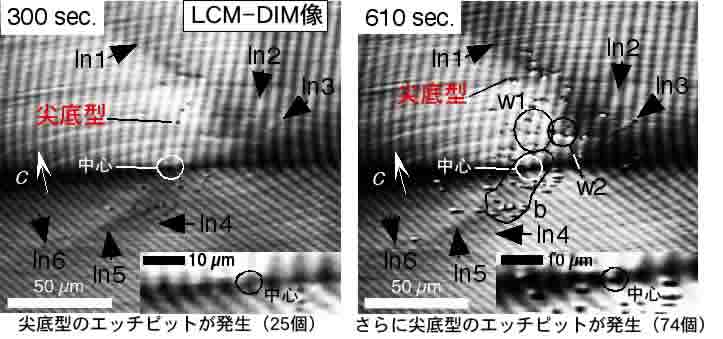
それでは次に,3)で観察された「線状」のコントラストが「転位」であることを証明します.そのために,観察セルの温度を結晶を成長させていた20℃から28℃へと上昇させ,結晶が次第に溶けてゆく様子をLCM-DIMを用いてその場観察しました.その様子を下図に示します.時間とともに,渦巻成長丘の中心付近では,底が尖って深い「尖底型」のエッチピットが多数現れてくることが分かります.また,下図の右下の挿入図は,渦巻成長丘のちょうど中心部分を拡大した像です.渦巻成長丘のちょうど中心部分には,渦巻ステップを供給しているらせん転位由来の尖底型のエッチピットがやはり出現することが確認できます.ここで注目していただきたいのは,上左図のln1-ln6で示した線状のコントラストちょうど端の部分に必ず尖底型のエッチピットが発生することです.このことより,これらの線状のコントラストは,視野面平行方向に発達した「転位」であることが証明されます.このように,LCM_DIMを用いて,成長しているタンパク質結晶中の転位のその場観察に初めて成功しました.X線トポグラフの場合には,分解能は数-数十μmですので,分解能が約2桁向上していることも重要な点です.
また下図より,渦巻成長丘の中心付近には,渦巻ステップの発生に寄与しない多くの転位が存在することが分かります.これらの多くは刃状転位であると考えられます.
右下の挿入図よりまた,らせん転位に相当する尖底型エッチピットが一番最後に遅れて現れることが分かります.このことは,転位周囲の歪エネルギーが,(らせん転位)<(刃状転位)であることを示しており,転位理論と一致しています.
(転位・インクルージョンのその場観察ページのトップへ)
5)結晶中に観察される2種類の格子欠陥:「転位とマイクロ欠陥」
4)で結晶表面のエッチング実験を行った後,さらに他の格子欠陥を探すために,温度を30℃に上昇させてさらにエッチング実験を行った結果を下図に示します.さらに大きな未飽和度(30℃)でエッチングを200秒行っても,転位の路頭部分に相当する尖底型エッチピットは新たに発生してきませんでした.このことより,4)での28℃でのエッチングにより,転位由来の尖底型エッチピットは全て現れきっていることが分かります.また,800秒エッチングすると,尖底型に加えて新たに底が平たく浅い「平底型」のエッチピットが現れてきました.これら平底型のエッチピットの起源は「マイクロ欠陥」と呼ばれ,結晶中の「空孔」および結晶中に取り込まれた「不純物」であることが,本同らの研究によって報告されています(H. Hondoh, T. Nakada, Jpn. J. Appl. Phys., 4. (2004) 4529; H. Hondoh, T. Nakada, J. Crystal Growth, 275 (2005) e1423).ここで重要なのは,平底型エッチピットの出現に必要な未飽和度が尖底型に比べて大きい点です.この結果は,格子欠陥周囲の歪みエネルギーが,(転位)>(マイクロ欠陥)であることを示しています.このことは,エッチピットより計算した格子欠陥の密度が,(転位:1.9x10^(-4) [1/μm^2])<(マイクロ欠陥:1.4x10^(-2) [1/μm^2])であることと一致しています.すなわち,転位は大きな歪みエネルギーを持ち大変不安定であるため,少ししか発生しませんが,マイクロ欠陥の歪みエネルギーは小さいために多数発生することになります.さて,どちらの格子欠陥が,結晶のX線回折特性により大きな悪さをするのでしょうね? 明らかにできる日が待ち遠しいです.

(転位・インクルージョンのその場観察ページのトップへ)
6)インクルージョンと転位の関係
さて,本頁の最後に,3)で示した転位の発生源が結晶中のインクルージョンであることを説明します.下図は,観察セルの温度を上げ下げし,リゾチーム結晶の「溶解(左下図)・成長(右下図)」を繰り返した様子を位相差顕微鏡で観察下像です.溶解時には転位が白いコントラストで観察されます.これらが転位であることは,左下図左上の挿入図において,尖底型のエッチピットが観察されていることからも分かります.この結晶を成長させると,右下図に示したように,転位のいくつかから渦巻成長丘が発達してくる様子が分かります.また,溶解・成長時ともに,図中の下部に「白い雲状」のコントラストが観察されます.これらは,結晶中に取り込まれているインクルージョンを示しています.ここで注目していただきたいことは,左下図において転位がインクルージョンの端っこの部分に多く集中して現れている点です.この結果は,インクルージョンが転位の発生源になっていることを示しています.すなわち,最後の図に示したポンチ絵のようになっていると考えられます.


(転位・インクルージョンのその場観察ページのトップへ)
To the top page of in-situ observation of protein crystallization; タンパク質結晶化過程のその場観察のトップページへ
To the top page; トップページへ,